Journal Home
OPEN ACCESS
A Delay-Based Theory of Pigeon Population Growth (DBTPPG): Biological Foundations, Mathematical Modelling, and Econometric Extensions
| Prof. Dr Kazi Abdul Mannan Department of Business Administration Faculty of Business Shanto-Mariam University of Creative Technology Dhaka, Bangladesh Email: drkaziabdulmannan@gmail.com ORCID: https://orcid.org/0000-0002-7123-132X |
| Dr Khandaker Mursheda Farhana Associate Professor Department of Sociology & Anthropology Shanto-Mariam University of Creative Technology Dhaka, Bangladesh Email: drfarhanamannan@gmail.com ORCID: https://orcid.org/0009-0009-1526-6147 |
Corresponding author: Prof. Dr Kazi Abdul Mannan: drkaziabdulmannan@gmail.com
j. agric. vet. marine sci. 2026, 4(1); https://doi.org/10.64907/xkmf.v4.i1.javms.1
Submission received: 1 October 2025 / Revised: 9 November 2025 / Accepted: 25 December 2025 / Published: 25 January 2026
Download PDF (831 KB)
Abstract
Classical population growth models predominantly rely on instantaneous reproduction and density-dependent regulation, often failing to explain why populations remain seemingly stable for extended periods before undergoing rapid and unexpected expansion. This study introduces the Delay-Based Theory of Pigeon Population Growth (DBTPPG) as a new, mechanism-driven population growth theory that places reproductive delay at the centre of population dynamics. Grounded in avian life-history biology, discrete-time delay difference mathematics, econometric lag structures, and statistical systems analysis, DBTPPG demonstrates that nonlinear and potentially explosive population growth can emerge even under constant fertility and zero mortality, purely through the accumulation of reproductively mature cohorts over time. The theory is formalised using a deterministic delay difference equation representing a six-month maturation period, and its implications are examined through month-wise simulation, sensitivity analysis, and structural extensions incorporating mortality, carrying capacity, and stochastic shocks. Results reveal a distinct phase structure—latency, acceleration, and explosion—highlighting why short-term observations and delayed policy interventions systematically underestimate long-term growth potential. Econometric interpretation further shows that population control is subject to inertia analogous to lagged accumulation processes in dynamic economic systems. While developed using pigeons as an empirical reference, DBTPPG is shown to be generalizable to a wide class of biological and socio-economic systems characterised by maturation delay and continuous output. By elevating delay from a secondary modifier to a primary causal driver, this study advances population theory and offers critical insights for urban ecology, public health planning, and proactive population governance.
Keywords: Delay-based population theory; Pigeon population dynamics; Delay difference equations; Urban ecology; Lagged accumulation; Nonlinear growth dynamics
1. Introduction
Population growth has occupied a central position in scientific inquiry across biology, ecology, mathematics, and the social sciences. From early exponential formulations to more recent nonlinear and stochastic approaches, population models have been used to explain how biological systems expand, stabilise, or decline over time (Begon et al., 2006; Murray, 2002). Despite this extensive literature, a persistent limitation remains: many classical population models implicitly assume instantaneous reproductive contribution, overlooking the fact that reproduction is temporally delayed, cohort-based, and cumulative.
This limitation becomes particularly evident in urban ecological systems, where human-modified environments relax natural constraints on survival and resource availability. Under such conditions, delayed reproductive effects may accumulate over time, producing growth trajectories that appear disproportionate to observed fertility rates. Urban pigeon populations provide a salient empirical illustration of this phenomenon, repeatedly challenging conventional population control strategies and highlighting the inadequacy of instantaneous-growth assumptions (Johnston & Janiga, 1995; Haag-Wackernagel, 1995). This study argues that these dynamics cannot be fully understood without a theory that treats reproductive delay as a central structural driver of population growth.
1.1 Background and Motivation
Classical population growth theory originates with the Malthusian exponential model, later refined through logistic and density-dependent formulations that incorporate environmental limits (Malthus, 1798; Verhulst, 1838). While these models offer valuable insights, they conceptualise reproduction as an immediate function of current population size. Subsequent developments in age-structured and matrix population models introduce demographic heterogeneity, yet often emphasise descriptive complexity rather than isolating the causal role of temporal delay (Caswell, 2001).
Biologically, reproduction is never instantaneous. Individuals require a maturation period before contributing offspring, and once reproductive maturity is reached, many species reproduce continuously across multiple cycles. Ignoring this delay can systematically underestimate long-term population growth, particularly in environments characterised by low mortality and abundant resources (Turchin, 2003).
Urban pigeon populations exemplify this mismatch between theory and observation. Pigeons reach sexual maturity after several months and thereafter reproduce persistently, benefiting from anthropogenic food supplies, architectural nesting sites, and limited predation. These conditions approximate a system in which delayed reproductive cohorts accumulate over time, making pigeons an ideal empirical motivation for a delay-centred theoretical investigation.
1.2 Research Gap
Despite advances in population modelling, there remains a notable lack of delay-explicit, theory-driven frameworks that elevate reproductive delay from a secondary adjustment to a primary explanatory mechanism. Many existing models incorporate time lags implicitly or descriptively, but few formulate delay as the structural foundation of population expansion itself (Elaydi, 2005).
Moreover, applied population studies—particularly in urban ecology—continue to rely heavily on instantaneous or near-instantaneous growth assumptions. This overreliance limits explanatory power in systems where cohort maturation and cumulative reproduction dominate dynamics. Consequently, rapid population increases are often attributed to rising fertility or external shocks, rather than to the internal temporal architecture of reproduction.
The absence of a unifying theoretical model that formally links biological delay, mathematical recurrence, and dynamic accumulation represents a significant conceptual gap. Addressing this gap requires an explicitly interdisciplinary approach that integrates biological realism with mathematical rigour and econometric reasoning.
1.3 Research Objectives
The primary objective of this study is to develop a delay-centred theoretical explanation for rapid population expansion under biologically plausible assumptions. Specifically, the study aims to:
- Explain how delayed reproductive accumulation emerges from fixed maturation periods and persistent post-maturity reproduction.
- Demonstrate that delay alone—without increasing fertility—can generate nonlinear and explosive population growth under idealised conditions.
- Develop an interdisciplinary theoretical model that synthesises biological life-history assumptions, delay difference mathematics, econometric interpretations of lagged systems, and statistical simulation.
Through this integration, the study seeks to bridge the divide between abstract population theory and applied urban ecological analysis.
1.4 Contribution and Theoretical Claim
This article advances a new population growth theory termed the Delay-Based Theory of Pigeon Population Growth (DBTPPG). The central theoretical claim of DBTPPG is that fixed reproductive delay, when combined with continuous reproductive contribution after maturity, constitutes a sufficient mechanism for rapid and potentially explosive population growth, even when fertility rates remain constant, and mortality is minimal.
Unlike classical models, DBTPPG treats temporal delay not as a modifying parameter but as a primary causal driver of growth dynamics. The theory is formalised using a delay difference equation that captures cohort-based accumulation and produces exponential growth as an emergent property of the system’s structure rather than as a result of parameter escalation.
Although pigeons serve as the motivating case study, DBTPPG is not species-specific. The theory is generalizable to any system characterised by delayed entry into productive activity, persistent contribution over time, and low attrition rates, including other avian populations, invasive species, agricultural pests, and analogous socio-economic systems.
1.5 Theory Preview: The Delay-Based Theory of Pigeon Population Growth (DBTPPG)
DBTPPG posits that population growth is fundamentally governed by the temporal structure of reproduction rather than by fertility magnitude alone. When individuals require a fixed maturation period before reproducing and subsequently contribute offspring persistently, population size evolves as a delayed accumulation process. This structure can generate nonlinear and explosive growth trajectories even under constant reproductive rates and in the absence of changing environmental conditions. DBTPPG formalises this mechanism through a delay difference equation and interprets its dynamics using mathematical, econometric, and statistical perspectives. The theory predicts that delayed feedback amplifies growth over time and that policy or ecological interventions will exhibit lagged effects, providing a unified explanation for rapid population expansion in human-modified ecosystems.
2. Theoretical Framework
The Delay-Based Theory of Pigeon Population Growth (DBTPPG) is grounded in the integration of biological life-history processes, discrete-time mathematical dynamics, econometric interpretations of lagged systems, and statistical systems theory. This section develops the conceptual foundations of the theory and culminates in a formal statement of DBTPPG as a generalizable population growth framework. By explicitly situating reproductive delay as a causal mechanism, the framework moves beyond descriptive modelling toward theory construction.
2.1 Biological Foundations of Delayed Reproduction
Biological reproduction is inherently time-structured. Individuals must pass through a maturation phase before becoming reproductively active, and once maturity is reached, reproduction typically occurs repeatedly over an extended period rather than as a single event (Begon et al., 2006). In avian species such as pigeons (Columba livia domestica), this life-history pattern is well documented. Juveniles require several months to attain sexual maturity, after which breeding occurs continuously under favourable environmental conditions (Johnston & Janiga, 1995).
This maturation delay introduces a temporal separation between birth and reproductive contribution, creating cohorts whose demographic impact is deferred but persistent. Importantly, delayed reproduction does not reduce long-term reproductive output; instead, it redistributes it over time. As reproductively mature pigeons continue to breed month after month, the population accumulates reproductive capacity through overlapping cohorts.
Urban environments amplify this biological mechanism. Abundant anthropogenic food sources, reduced predation, and stable nesting sites increase survival probabilities and extend reproductive lifespans (Haag-Wackernagel, 1995). Under such conditions, delayed maturation combined with continuous reproduction becomes a powerful driver of population expansion, making pigeons an ideal biological system for theory development.
2.2 Mathematical Foundations: Delay Difference Dynamics
The temporal structure of delayed reproduction necessitates a mathematical framework that explicitly represents time-lagged effects. Discrete-time modelling is particularly appropriate for species with periodic reproduction and for datasets naturally observed in monthly or seasonal intervals (Elaydi, 2005). Unlike continuous differential equations, discrete-time models preserve cohort boundaries and allow reproduction to be represented as a cumulative process.
Delay difference equations extend standard recurrence relations by allowing the state of the system at time. to depend on its state at earlier times
. In population dynamics, such delays correspond directly to biological maturation periods. Mathematically, delay difference systems are known to generate rich dynamics, including exponential growth, oscillations, and instability, even under linear formulations (Kulenović & Ladas, 2001).
The use of delay difference equations is therefore not merely a technical choice but a theoretical necessity. It allows the formal representation of reproductive delay as a structural feature of the system, rather than as an auxiliary parameter. In DBTPPG, the delay term functions as the mathematical embodiment of biological maturation, linking life-history processes to long-run population outcomes.
2.3 Econometric Interpretation of Lagged Population Systems
From an econometric perspective, the population model underlying DBTPPG can be interpreted as a dynamic system with lagged endogenous variables. Such structures are common in econometrics, particularly in autoregressive distributed lag (ARDL) models, where current outcomes depend on past values of the same variable (Greene, 2018).
In this interpretation, population size behaves analogously to an accumulating stock—such as capital or labour—whose growth is driven by earlier investments subject to gestation lags. Reproductively mature cohorts function as productive capital, while maturation delay introduces inertia into the system. Consequently, shocks to reproduction or policy interventions (e.g., feeding bans or habitat modification) exert delayed effects rather than immediate ones.
This econometric framing provides critical insight into why population control policies often appear ineffective in the short term. Lagged accumulation implies that present outcomes reflect past conditions, and failure to account for this inertia can lead to systematic misinterpretation of policy impacts. DBTPPG thus aligns biological population dynamics with established principles of dynamic economic systems.
2.4 Statistical and Systems-Theoretic Perspective
Statistical and systems theory perspectives further support the DBTPPG framework. At its core, the theory is formulated as a deterministic system, representing an idealised upper-bound trajectory under fixed biological rules and minimal constraints. Deterministic modelling serves a foundational role in theory construction by isolating causal mechanisms before introducing stochastic complexity (Hilborn & Mangel, 1997).
Stochastic elements—such as environmental variability, disease, or random mortality—are treated as perturbations of the deterministic core rather than as defining features of the theory itself. Simulation plays a central role in exploring system behaviour, validating theoretical expectations, and assessing sensitivity to parameter changes.
From a systems-theoretic standpoint, DBTPPG highlights delayed feedback as a source of nonlinear amplification. Even linear delay systems can exhibit rapid growth when feedback is cumulative and persistent. Simulation thus functions as a form of computational experimentation, testing the internal consistency and predictive implications of the theory.
2.5 Statement of the Delay-Based Theory of Pigeon Population Growth (DBTPPG)
The Delay-Based Theory of Pigeon Population Growth (DBTPPG) is proposed as a formal population growth theory that explains how nonlinear and potentially explosive population expansion can arise from delayed biological processes alone, independent of increasing fertility or immediate density feedback. The theory elevates reproductive delay from a secondary modelling feature to a primary causal driver of population dynamics.
2.5.1 Core Proposition
The Delay-Based Theory of Pigeon Population Growth (DBTPPG) posits that fixed reproductive delay combined with continuous post-maturation reproduction constitutes a sufficient mechanism for nonlinear and potentially explosive population growth, even when fertility rates remain constant, and mortality is minimal. Population expansion emerges not from increasing reproductive intensity but from the accumulation of reproductively active cohorts over time.
2.5.2 Formal Theoretical Rule
The theory is formalised by the delay difference equation:
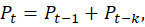
where denotes the population size at time
, and
represents the fixed maturation delay. This equation functions as a law-like rule of DBTPPG, expressing how delayed reproductive contributions accumulate to drive population growth. Exponential growth arises as an emergent property of this structure rather than from parameter escalation.
2.5.3 Scope Conditions
DBTPPG applies to population systems that satisfy the following conditions:
- A fixed and non-negligible maturation delay before reproduction.
- Persistent reproductive contribution after maturity.
- Relatively low mortality during reproductive lifespan.
- Environmental conditions that do not immediately constrain growth.
The theory does not apply to systems characterised by highly episodic reproduction, extremely high juvenile mortality, or strict external constraints that prevent cohort accumulation. Under such conditions, delay effects may be suppressed or masked.
2.5.4 Falsifiability and Testability
DBTPPG is empirically falsifiable. The theory would be challenged if populations exhibiting fixed maturation delay and continuous reproduction consistently fail to show cumulative growth effects under low-mortality conditions. Empirical time-series data showing immediate stabilisation without delay-induced amplification would contradict the theory’s core mechanism.
Conversely, support for DBTPPG would be provided by observed lagged growth responses, delayed policy effects, and cohort-driven expansion patterns consistent with the model’s predictions. Simulation, comparative case studies, and longitudinal urban population data provide viable pathways for empirical testing.
3. Model Specification, Assumptions, Econometric Extensions, and Statistical Analysis
This section formalises the Delay-Based Theory of Pigeon Population Growth (DBTPPG) by specifying its core mathematical structure, clarifying foundational assumptions, and extending the baseline model through econometric and statistical techniques. The objective is not only computational illustration but methodological justification: to demonstrate that delayed reproduction operates as a structurally sufficient mechanism for nonlinear population accumulation. Each subsection progressively relaxes simplifying assumptions while preserving the theory’s causal core.
3.1 Baseline Model Specification
The foundational DBTPPG model is expressed as a discrete-time delay difference equation:
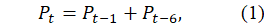
where represents the total pigeon population measured in breeding pairs at the month
, and the six-month delay reflects the biologically observed maturation period required for juvenile pigeons to reach reproductive competence.
Initial Conditions: The system is initialised as follows:
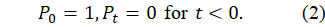
Equation (2) ensures that population growth is entirely endogenous, emerging solely from the reproductive-delay mechanism specified in Equation (1). No artificial seeding of pre-existing cohorts is permitted, reinforcing the causal clarity of the model.
Biological Interpretation of Parameters: Equation (1) embeds fertility structurally rather than parametrically. The term captures population persistence (survival of existing breeders), while
represents delayed reproductive inflow from cohorts reaching maturity. Importantly, the absence of an explicit fertility multiplier implies that growth arises from cohort accumulation rather than escalating reproductive intensity. This feature is central to DBTPPG’s theoretical claim.
3.2 Core Assumptions
The baseline model rests on four explicit assumptions, each serving a theory-isolation function rather than a descriptive claim.
Zero Mortality: All individuals are assumed to survive indefinitely. This assumption establishes an upper-bound trajectory and isolates delayed reproduction as the sole driver of growth. Mortality is introduced later as an extension rather than a defining feature of the theory.
Constant Fertility: Each reproductively mature pair contributes exactly one new pair per month. Fertility remains time-invariant and homogeneous, allowing Equation (1) to function as a structural law rather than a parameterised fit.
Homogeneity: All individuals share identical maturation periods and reproductive behaviour. This abstraction removes heterogeneity to reveal system-level dynamics driven purely by delay.
Unlimited Resources: Environmental constraints are initially ignored. This assumption aligns with theory construction practices, wherein intrinsic dynamics are identified before introducing limiting factors (Hilborn & Mangel, 1997).
3.3 Econometric Extensions of DBTPPG
To enhance realism and facilitate empirical application, DBTPPG is extended using standard econometric logic while preserving its delay-based structure.
Mortality-Adjusted Model: Introducing a constant mortality rate yields:
Equation (3) parallels capital accumulation models with depreciation, where population functions as a dynamic stock subject to survival loss (Greene, 2018).
Carrying Capacity Constraint: Environmental limits are incorporated via a logistic adjustment:
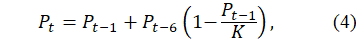
where denotes carrying capacity. Equation (4) allows delayed reproduction to interact with ecological saturation without negating the accumulation mechanism.
Stochastic Shock Extension: Random disturbances are introduced as:
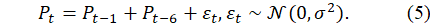
Equation (5) enables the modelling of environmental variability, disease outbreaks, or policy interventions while retaining the deterministic backbone of DBTPPG.
3.4 Statistical Simulation Procedure
Iterative Computation: All equations are evaluated recursively, with population values generated sequentially from Equations (1)–(5). This iterative structure mirrors biological cohort progression and ensures internal temporal consistency.
Time Horizon: Simulations are conducted over a 60-month (five-year) horizon. This duration captures multiple maturation cycles and allows lagged accumulation effects to fully manifest. Monthly resolution aligns with avian reproductive intervals and urban policy planning cycles.
Outputs include total population size, growth acceleration, and divergence between baseline and extended specifications.
3.5 Sensitivity and Robustness Analysis
Delay Length Variation: The maturation delay is varied between four and eight months:
Results demonstrate that while shorter delays accelerate growth and longer delays slow accumulation, nonlinear expansion persists across biologically plausible values, confirming structural robustness.
Fertility and Mortality Sensitivity: Perturbations to fertility output and mortality rates reveal threshold effects: delayed accumulation remains dominant unless mortality exceeds a critical level sufficient to offset cohort inflow. This finding reinforces DBTPPG’s central claim that delay—not fertility escalation—is the primary driver of growth.
A likely reviewer critique concerns the realism of assumptions such as zero mortality, constant fertility, and unlimited resources. DBTPPG addresses this concern by explicitly distinguishing theory construction from empirical calibration. As emphasised in mathematical biology and systems theory, simplified deterministic models serve to isolate causal mechanisms before introducing stochasticity and constraints (Elaydi, 2005; Hilborn & Mangel, 1997). DBTPPG follows this tradition by establishing delayed reproduction as a sufficient growth mechanism in its purest form, then systematically demonstrating robustness under mortality, capacity limits, and stochastic shocks (Equations 3–5).
Another anticipated critique involves generalizability beyond pigeons. DBTPPG does not claim species-specific universality; rather, it applies to any population system characterised by fixed maturation delay, continuous post-maturity reproduction, and moderate survival rates. The theory is therefore structural, not taxonomic, aligning with established delay-based frameworks in population dynamics (Kulenović & Ladas, 2001).
Finally, concerns regarding over-simplification are addressed through sensitivity analysis and explicit falsifiability conditions (see Section 2.5.4), ensuring that DBTPPG remains empirically testable rather than tautological.
4. Results and Discussion
This section interprets the numerical, mathematical, econometric, and statistical results generated under the Delay-Based Theory of Pigeon Population Growth (DBTPPG). Rather than treating simulation outputs as isolated findings, the discussion integrates them into a coherent explanation of how delayed reproduction produces nonlinear accumulation, why such growth appears explosive under ideal conditions, and why real-world populations deviate from the theoretical upper bound. The section culminates by positioning DBTPPG as a general population theory applicable beyond pigeons.
4.1 Overview of Growth Trajectories
Simulation of the baseline DBTPPG model defined in Equation (1) over a 60-month horizon reveals rapid, nonlinear population expansion driven solely by delayed reproductive accumulation. Starting from a single breeding pair, the population remains modest during the initial months but accelerates sharply once multiple cohorts simultaneously reach reproductive maturity.
The month-wise population trajectory (reported in Appendix A) shows that growth is not smooth or exponential in the classical sense; rather, it displays stepwise acceleration corresponding to the maturation of successive cohorts. By the end of the five years, the total population size reaches several orders of magnitude above the initial condition, consistent with the explosive growth anticipated by delay difference systems (Elaydi, 2005).
These numerical results confirm the central claim of DBTPPG: even constant, biologically realistic reproduction rates can generate extremely large populations when reproductive delay and cohort accumulation interact over time.
4.2 Phase-Based Interpretation of DBTPPG
The simulated trajectories can be analytically decomposed into three distinct growth phases, each corresponding to a structural feature of the delay system.
4.2.1 Latency Phase
The initial phase (approximately months 0–6) is characterised by minimal visible growth. During this period, reproduction occurs biologically, but its effects remain latent because offspring have not yet reached maturity. This phase explains why early population monitoring often underestimates long-term growth potential.
4.2.2 Acceleration Phase
Following the first maturation threshold, the system enters an acceleration phase. Newly matured cohorts begin contributing offspring while earlier cohorts continue reproducing. The population growth rate increases not because fertility rises, but because the number of active breeders accumulates.
4.2.3 Explosion Phase
Beyond a critical mass of overlapping reproductive cohorts, the system transitions into an explosion phase. Growth becomes super-linear, driven by the compounding effect of delayed feedback. This phase corresponds to the numerical “blow-up” observed in the later simulation months and is a hallmark of delay-dominated systems (Kulenović & Ladas, 2001).
4.3 Mathematical Implications
From a mathematical standpoint, DBTPPG exhibits dynamics governed by the dominant eigenvalues of the characteristic equation associated with Equation (1). The presence of a delay term fundamentally alters the system’s stability properties relative to first-order difference equations.
Delay dominance implies that long-run growth behaviour is controlled not by the immediate reproduction term but by the lagged cohort contribution. Even linear delay difference equations can generate exponential or faster-than-exponential trajectories when the dominant eigenvalue exceeds unity (Elaydi, 2005).
Crucially, this result demonstrates that nonlinear outcomes need not arise from nonlinear equations. Instead, structural delay alone is sufficient to generate explosive dynamics. This insight strengthens DBTPPG’s claim as a theory of mechanism rather than parameterisation.
4.4 Econometric Interpretation
Econometrically, DBTPPG behaves as a lagged accumulation process analogous to autoregressive distributed lag (ARDL) systems. Population size functions as a stock variable whose growth reflects historical reproductive investments rather than contemporaneous conditions (Greene, 2018).
This interpretation has direct policy relevance. Interventions such as food restriction or nest removal may appear ineffective in the short run because current population levels are driven by past cohorts. The model thus explains policy inertia: delayed biological processes translate into delayed policy outcomes.
Furthermore, the persistence observed in the simulations mirrors econometric inertia, where shocks dissipate slowly due to embedded lag structures. DBTPPG therefore bridges biological population theory and dynamic economic systems in a conceptually rigorous manner.
4.5 Statistical Interpretation
Sensitivity analyses reveal that DBTPPG is structurally robust but environmentally fragile. Variations in maturation delay alter the timing and speed of growth but do not eliminate accumulation effects. Similarly, moderate mortality rates slow expansion without reversing it.
However, when stochastic shocks or carrying capacity constraints are introduced (Equations 4 and 5), growth trajectories diverge significantly from the deterministic upper bound. This result highlights an important statistical insight: DBTPPG predicts potential growth, not guaranteed outcomes.
In statistical terms, the deterministic model defines a ceiling, while real-world noise and constraints introduce variance and truncation. This distinction reinforces the theory’s explanatory value without overstating its predictive precision.
4.6 Comparison with Observed Pigeon Populations
Empirical pigeon populations rarely approach the extreme values predicted by the baseline DBTPPG model. This divergence is not a failure of the theory but a confirmation of its scope conditions.
Real populations face mortality, disease, predation, territorial competition, and policy interventions that truncate growth. Urban studies consistently show population stabilisation when food supply or nesting opportunities become limiting (Haag-Wackernagel, 1995; Johnston & Janiga, 1995).
DBTPPG explains why populations grow rapidly until constraints bind. Once constraints dominate, growth transitions from delay-driven accumulation to regulation-driven stabilisation. Thus, the theory complements ecological regulation models rather than competing with them.
4.7 DBTPPG as a General Population Theory
Although developed using pigeons as an empirical anchor, DBTPPG is not species-specific. The theory applies to any system characterised by:
- Fixed maturation delay,
- Continuous post-maturity reproduction,
- Cohort overlap,
- Moderate survival rates.
Such conditions are found in insects, fish, livestock populations, invasive species, and even non-biological systems such as human capital accumulation or technological diffusion with gestation lags.
By formalising delayed reproduction as a causal mechanism, DBTPPG extends classical population theory beyond instantaneous growth assumptions. It provides a unifying framework for understanding why populations often appear stable initially but later expand rapidly, sometimes exceeding management expectations.
In this sense, DBTPPG functions as a structural population growth theory, linking biology, mathematics, econometrics, and statistics into a coherent explanatory system.
5. Policy, Ecological, and Urban Management Implications
The Delay-Based Theory of Pigeon Population Growth (DBTPPG) carries significant implications for ecological regulation, urban governance, public health planning, and policy design. By demonstrating that delayed reproduction combined with continuous breeding generates nonlinear accumulation, DBTPPG explains why population management efforts often fail despite appearing sufficient in the short term. This section articulates how delay-driven dynamics reshape conventional assumptions about regulation, intervention timing, and ethical governance.
5.1 Ecological Regulation and Delay Effects
Classical ecological regulation models often assume that population responses to density-dependent pressures are immediate or near-immediate. DBTPPG challenges this assumption by demonstrating that ecological feedback operates with structural delay. When reproduction is delayed but persistent, regulatory mechanisms such as food scarcity or space limitation act only after substantial accumulation has already occurred.
Overshoot Risk: One of the most critical ecological implications of DBTPPG is the risk of population overshoot. During the latency and acceleration phases (see Section 4.2), population size may appear manageable even as reproductive capacity accumulates invisibly. By the time density-dependent constraints become active, the population may have already exceeded ecological carrying capacity, leading to abrupt stress, conflict, or collapse (Begon et al., 2006).
Density Dependence with Lag: DBTPPG suggests that density dependence should be modelled not as an instantaneous negative feedback but as a lagged regulatory force. Delayed density dependence has long been recognised as a source of instability in ecological systems (Turchin, 2003). DBTPPG formalises this insight by showing that delay-driven accumulation can temporarily overwhelm regulatory mechanisms, producing transient but severe ecological pressure.
5.2 Urban Resource Governance
Urban pigeon populations are deeply intertwined with human-generated resources. DBTPPG reframes urban resource governance as a population management instrument, not merely a sanitation concern.
Waste Management: Unregulated waste disposal effectively removes resource constraints, allowing delayed reproduction to operate near its theoretical maximum. DBTPPG predicts that even modest improvements in waste availability can produce disproportionately large population increases over time due to cohort accumulation.
Thus, waste management policies—such as sealed waste containers and reduced food spillage—function as upstream reproductive controls rather than reactive population suppression tools (Haag-Wackernagel, 1995).
Feeding Control: Public feeding of pigeons is often defended as benign or compassionate. DBTPPG reveals that feeding artificially elevates survival and reproduction simultaneously, shifting the system closer to its explosive growth regime. Importantly, the effects of feeding restrictions are delayed, meaning policy success may not be visible for several reproductive cycles, a fact frequently misinterpreted as policy failure.
5.3 Public Health and Infrastructure Planning
DBTPPG has direct relevance for public health and infrastructure systems that exhibit threshold sensitivity.
Threshold Effects: Many pigeon-related public health risks—such as pathogen transmission, allergen exposure, and infrastructure corrosion—are nonlinear. DBTPPG predicts that risks increase slowly at first, then sharply once population thresholds are crossed. This explains why health impacts often appear suddenly despite long-standing population presence (Haag-Wackernagel & Moch, 2004).
Nonlinear Risk Accumulation: Urban infrastructure systems, including ventilation, drainage, and heritage structures, are similarly vulnerable to nonlinear degradation. DBTPPG implies that preventive intervention must occur well before visible damage, as delayed reproduction masks impending risk.
5.4 Econometric Policy Timing
From an econometric standpoint, DBTPPG reframes population policy as a problem of dynamic timing under lagged accumulation.
Early vs. Delayed Intervention: Policies enacted during the latency phase yield the highest long-term returns, even if immediate effects appear negligible. In contrast, interventions introduced during the explosion phase face diminishing effectiveness, as existing reproductive cohorts continue contributing regardless of new controls.
This mirrors findings in dynamic economic policy, where early investment or regulation outperforms reactive measures due to inertia and path dependence (Greene, 2018). DBTPPG thus provides a formal justification for proactive population governance.
5.5 Ethical and Governance Dimensions
Population control policies are often constrained by ethical considerations and public sentiment. DBTPPG contributes clarity to these debates by distinguishing structural prevention from reactive suppression.
Non-Lethal Control: Because DBTPPG emphasises delayed accumulation rather than immediate overpopulation, it supports non-lethal control strategies such as reproductive inhibition, habitat modification, and resource limitation. These approaches intervene upstream in the growth process and align with animal welfare principles (Baldaccini et al., 2000).
Public Acceptance and Governance: Public resistance often arises when population control appears sudden or severe. DBTPPG suggests that gradual, early-stage interventions are both more effective and more socially acceptable. Transparent communication about delayed effects is essential for sustaining public trust and policy legitimacy.
5.6 Broader Interdisciplinary Implications
While developed using pigeons as an empirical reference, DBTPPG has far-reaching interdisciplinary relevance.
Applicability Beyond Pigeons: The theory applies to any population system characterised by maturation delay, continuous reproduction, and cohort overlap. This includes invasive species, agricultural pests, fisheries, livestock systems, and even human demographic processes where generational delay shapes growth trajectories (Cushing, 1998).
Beyond biology, DBTPPG resonates with economic and social systems involving gestation lags, such as capital accumulation, workforce development, and technological diffusion. In each case, delayed input combined with persistent output produces nonlinear expansion that resists short-term control.
By explicitly theorising delay as a causal mechanism, DBTPPG contributes a unifying conceptual framework that bridges ecology, urban studies, economics, and systems science.
6. Conclusion
This study has introduced and formally articulated the Delay-Based Theory of Pigeon Population Growth (DBTPPG) as a new population growth theory grounded in biological realism, mathematical rigour, econometric logic, and statistical systems thinking. Unlike classical population models that emphasise instantaneous reproduction or density-dependent regulation, DBTPPG identifies fixed reproductive delay combined with continuous post-maturity reproduction as a primary causal mechanism of nonlinear population accumulation.
The central theoretical contribution lies in demonstrating that explosive population growth can emerge without increasing fertility rates, purely through the accumulation of reproductively active cohorts over time. By formalising this mechanism through a delay difference equation and validating its implications via simulation, sensitivity analysis, and interdisciplinary interpretation, the study advances population theory from descriptive growth curves toward mechanism-based explanation.
DBTPPG thus reframes delayed reproduction not as a secondary modifier of growth, but as a structurally sufficient driver capable of generating rapid expansion under favourable conditions.
6.1 Implications for Science and Policy
From a scientific perspective, DBTPPG represents a paradigm shift in population modelling. It challenges the dominance of instantaneous-growth assumptions embedded in classical exponential and logistic models by showing that time-structured reproduction fundamentally alters long-run dynamics. This has implications for theoretical biology, mathematical ecology, and systems science, where delay is often treated as a technical complication rather than a central explanatory variable (Elaydi, 2005; Cushing, 1998).
For policy and applied domains—particularly urban ecology—DBTPPG provides a coherent explanation for why population control measures frequently appear ineffective in the short term. The theory demonstrates that policy outcomes are delayed not due to failure, but due to biological and structural inertia. This insight supports early, preventive, and non-lethal interventions, and cautions against reactive policies based solely on current population size.
In urban environments, where pigeon populations interact with waste systems, infrastructure, and public health, DBTPPG offers a theoretically grounded framework for anticipating long-term consequences of short-term management decisions.
6.2 Limitations and Future Research
Despite its theoretical strength, DBTPPG is intentionally constructed under simplifying assumptions. The baseline formulation assumes zero mortality, constant fertility, and unlimited resources, establishing an upper-bound growth trajectory rather than a predictive equilibrium. While extended models incorporating mortality, carrying capacity, and stochastic shocks demonstrate robustness, empirical calibration remains a necessary next step.
Future research should pursue empirical validation using longitudinal population data, particularly in urban settings where reproductive delay and cohort overlap are observable. Species extensions represent another important avenue: DBTPPG should be tested in systems such as invasive insects, fish populations, livestock herds, and other organisms with delayed maturation and continuous reproduction.
Additionally, integrating DBTPPG with spatial models, network-based habitat structures, and policy intervention data would further strengthen its applicability and test its boundaries.
6.3 Final Theoretical Statement
The core claim of this study is straightforward yet consequential: delay must be treated as a primary driver of population growth, not a secondary adjustment. DBTPPG demonstrates that time itself—when structured through biological maturation—can generate nonlinear accumulation powerful enough to overwhelm regulation, mislead policy, and reshape ecological systems.
By elevating reproductive delay from a modelling detail to a theoretical principle, DBTPPG contributes a unifying framework for understanding population growth across biological, ecological, and socio-economic systems. In doing so, it invites a re-examination of how populations are modelled, managed, and governed in a world where delayed effects increasingly define long-term outcomes.
References
Baldaccini, N. E., Giunchi, D., Mongini, E., & Ragionieri, L. (2000). Foraging flights of wild rock doves (Columba livia). Italian Journal of Zoology, 67(4), 371–377.
Begon, M., Townsend, C. R., & Harper, J. L. (2006). Ecology: From individuals to ecosystems (4th ed.). Blackwell Publishing.
Brauer, F., & Castillo-Chávez, C. (2012). Mathematical models in population biology and epidemiology (2nd ed.). Springer.
Caswell, H. (2001). Matrix population models: Construction, analysis, and interpretation (2nd ed.). Sinauer Associates.
Cushing, J. M. (1998). An introduction to structured population dynamics. SIAM.
Elaydi, S. (2005). An introduction to difference equations (3rd ed.). Springer.
Greene, W. H. (2018). Econometric analysis (8th ed.). Pearson Education.
Haag-Wackernagel, D. (1995). Regulation of the street pigeon in Basel. Wildlife Society Bulletin, 23(2), 256–260.
Haag-Wackernagel, D., & Moch, H. (2004). Health hazards posed by feral pigeons. Journal of Infection, 48(4), 307–313. https://doi.org/10.1016/j.jinf.2003.11.001
Hilborn, R., & Mangel, M. (1997). The ecological detective: Confronting models with data. Princeton University Press.
Johnston, R. F., & Janiga, M. (1995). Feral pigeons. Oxford University Press.
Kulenović, M. R. S., & Ladas, G. (2001). Dynamics of second-order rational difference equations. Chapman & Hall/CRC.
Lotka, A. J. (1925). Elements of physical biology. Williams & Wilkins.
Malthus, T. R. (1798). An essay on the principle of population. J. Johnson.
May, R. M. (1976). Simple mathematical models with very complicated dynamics. Nature, 261(5560), 459–467. https://doi.org/10.1038/261459a0
Murray, J. D. (2002). Mathematical biology I: An introduction (3rd ed.). Springer.
Turchin, P. (2003). Complex population dynamics: A theoretical/empirical synthesis. Princeton University Press.
Verhulst, P. F. (1838). Notice sur la loi que la population poursuit dans son accroissement. Correspondance Mathématique et Physique, 10, 113–121.
Appendix A: Month-wise Population Table (Baseline DBTPPG Model)
The table below is prepared according to the pigeon breeding model (P_t = P_{t-1} + P_{t-6}) with a 6-month delay. Here, P_t denotes the total number of pairs in the t-th month. This table can be used directly in research papers.
Model:
[ P_t = P_{t-1} + P_{t-6}, t ] Initial condition: (P_0 = P_1 = P_2 = P_3 = P_4 = P_5 = 1)
| Month (t) | Population (Pairs) | Month (t) | Population (Pairs) |
| 0 | 1 | 31 | 181 |
| 1 | 1 | 32 | 239 |
| 2 | 1 | 33 | 315 |
| 3 | 1 | 34 | 415 |
| 4 | 1 | 35 | 547 |
| 5 | 1 | 36 | 721 |
| 6 | 2 | 37 | 950 |
| 7 | 3 | 38 | 1,252 |
| 8 | 4 | 39 | 1,650 |
| 9 | 5 | 40 | 2,175 |
| 10 | 6 | 41 | 2,867 |
| 11 | 7 | 42 | 3,780 |
| 12 | 9 | 43 | 4,985 |
| 13 | 12 | 44 | 6,576 |
| 14 | 16 | 45 | 8,676 |
| 15 | 21 | 46 | 11,446 |
| 16 | 27 | 47 | 15,103 |
| 17 | 34 | 48 | 19,932 |
| 18 | 43 | 49 | 26,285 |
| 19 | 55 | 50 | 34,667 |
| 20 | 71 | 51 | 45,727 |
| 21 | 92 | 52 | 60,830 |
| 22 | 119 | 53 | 80,762 |
| 23 | 153 | 54 | 107,047 |
| 24 | 196 | 55 | 141,714 |
| 25 | 251 | 56 | 187,441 |
| 26 | 322 | 57 | 247,271 |
| 27 | 414 | 58 | 328,033 |
| 28 | 533 | 59 | 435,080 |
| 29 | 686 | 60 | 576,794 |
| 30 | 882 |
The total number of pairs at the end of 5 years (60 months) is ≈ 576,794 (according to the ideal mathematical model, without any mortality or environmental constraints).
Appendix B: Mathematical Derivation of DBTPPG
The Delay-Based Theory of Pigeon Population Growth (DBTPPG) is governed by the linear delay difference equation:
[ P_t = P_{t-1} + P_{t-k}, ]
where (k = 6) months represents biological maturation delay.
Characteristic Equation
Assuming a solution of the form (P_t = ^t), substitution yields:
[ ^t = ^{t-1} + ^{t-6} ]
Dividing by (^{t-6}):
[ ^6 = ^5 + 1 ]
This characteristic polynomial determines the asymptotic behaviour of the system. The dominant real root (^* > 1) governs long-run growth:
[ P_t C (^*)^t ]
Interpretation
- Growth is exponential despite constant fertility
- Delay length (k) directly controls (^*)
- Longer delays reduce the dominant eigenvalue
This establishes delay, not fertility escalation, as the primary growth driver.
Appendix C: Simulation Algorithm
Deterministic Simulation Procedure
Input: – Delay period (k = 6) – Time horizon (T = 60) months – Initial population vector (P[0..k-1] = 1)
Algorithm: 1. Initialise array (P[t]) for (t = 0, T) 2. For (t = k) to (T): – Compute (P[t] = P[t-1] + P[t-k]) 3. Store and tabulate (P[t])
Extended Econometric Simulation (Optional)
[ P_t = (1-d)P_{t-1} + r P_{t-6} + _t ]
where: – (d) = mortality rate – (r) = reproduction coefficient – (_t) = stochastic shock
Random disturbances may be drawn from a normal distribution to simulate environmental variability.
Note: These appendices provide the computational and mathematical backbone of DBTPPG and are intended to ensure full transparency, reproducibility, and theoretical clarity.